

Abstract
Visual perceptual learning (VPL) is long-term performance increase resulting from visual perceptual experience. Task-relevant VPL of a feature results from training of a task on the feature relevant to the task. Task-irrelevant VPL arises as a result of exposure to the feature irrelevant to the trained task. There are at least two serious problems. First, which stage of information processing is changed in association with task-relevant VPL is controversial. Second, no model has ever explained both task-relevant and task-irrelevant VPL. Here we propose a dual plasticity model, in which there are feature-based plasticity that is a change in a representation of the learned feature and task-based plasticity that is a change in processing of the trained task. While the two types of plasticity underlie task-relevant VPL, only feature-based plasticity lies under task-irrelevant VPL. This model provides a new comprehensive framework in which apparently contradictory results could be explained.
Keywords: Location specificity, Early visual cortex, Transfer, Reweighting, Task-relevant perceptual learning, Task-irrelevant perceptual learning
Introduction
It is well known that for the first several months after the birth, called the critical period, the early visual cortex goes through a major rewiring, which leads the infant to acquire important visual abilities (Hensch 2005a, Hensch 2005b, Hubel & Wiesel 1964, Morishita & Hensch 2008). However, this does not indicate that after the critical period, the visual system does not have the ability to learn or that the visual plasticity brake (inhibition on plasticity) is completely on. A huge amount of research results indicate that as a result of visual experiences, an adult can acquire a new skill of improving performance on a visual task and of maintaining the skill for months or even years (Fahle & Poggio 2002, Fine & Jacobs 2002, Gilbert & Li 2012, Gold & Watanabe 2010, Goldstone 1998, Karmarkar & Dan 2006, Lu et al 2011, Sagi 2011, Sagi & Tanne 1994, Sasaki & Watanabe 2012, Sasaki et al 2010, Seitz & Dinse 2007). For example, it is extremely difficult to discriminate the sex of a chick. However, as a result of training, an adult can learn to do this extremely difficult task fairly precisely (Eagleman 2011). In most cases a radiologist can easily find cancer that untrained observers cannot possibly do in an x-ray picture. These examples indicate that it is possible to acquire some visual skills long time after the critical period. The process and result of acquiring such visual skills is called visual perceptual learning (VPL). VPL is regarded as an important tool that can help clarify the basic mechanisms of visual and brain plasticity.
At the same time, training on a visual task can significantly strengthen the visual abilities of adult patients with amblyopia and other forms of abnormal vision, which is regarded as an example of a highly successful translation of basic research findings into practical and powerful real-world applications (Andersen et al 2010, Bennett et al 2001, Levi & Li 2009a, Li et al 2013, Ooi et al 2013, Polat et al 2004, Shibata & Watanabe 2012). In addition, it has been found that training older people on a visual task leads to better performance (Andersen et al 2010, Bower et al 2013, Deloss et al 2013). This raises the possibility that visual training can restore or at least improve a variety of visual abilities in older people that have declined with aging (Andersen 2012).
Thus, a clearer understanding of the mechanisms of VPL in adults should lead to new innovations in basic science and clinical intervention. For these reasons, research on VPL has become increasingly popular, as evidenced by the fact that around 150 papers on VPL were published in 2013 alone.
In spite of the hope that a large volume of data accumulation will lead to clarification of the mechanism of VPL, which in turn would lead to developing highly effective ways to greatly restore or improve damaged or declined vision, there has been a general tendency that is opposite to the hope. In reality, instead of having different models being converged to a unified view, several issues that highly relate to core mechanisms of VPL are becoming incrementally more controversial.
This review focuses on VPL of relatively primitive visual features including orientation, motion direction and luminance contrast. Visual processing of a primitive feature has been better clarified than most of other types of brain processing. Thus, better understanding of the basic mechanism of learning of a primitive visual feature may lead to better understanding of learning of other sensory modalities, learning of more complex visual processing including visual categorization and of more complex features such as objects and faces. The better understanding may also significantly contribute to development of clinical methods to improve and/or restore visual abilities that have been damaged by diseases or declined with aging. However, there are important differences between VPL of a primitive visual feature and these aspects of learning. Thus, interested readers should also read reviews on specialized fields. There have been excellent reviews and papers on learning of other sensory modalities (Beste & Dinse 2013, de Villers-Sidani & Merzenich 2011), learning of more complex aspects (Ashby & Maddox 2011, Op de Beeck & Baker 2010), learning of faces (Bi et al 2014), clinical applications of VPL (Levi 2012, Levi & Li 2009b), and effects of aging on VPL (Andersen 2012).
As there are active and passive natures in most types of learning and memory, VPL is classified into 2 types, task-relevant VPL and task-irrelevant VPL. Task-relevant VPL is defined as VPL of a feature that is relevant to a given task during training. Task-irrelevant VPL is defined as VPL of a feature that is irrelevant to a given task (Seitz & Watanabe 2005). We will review research on task-relevant VPL and then on task-irrelevant VPL.
Task-Relevant Visual Perceptual Learning
One of the most controversial issues in task-relevant VPL is which stage/brain area of visual information processing is changed in association with task-relevant VPL. According to which stage/area is assumed to be associated with task-relevant VPL, most models of task-relevant VPL are roughly classified as 3 different models: the early-stage model, mid-stage model, and late-stage model. Here we will review and discuss each model. “Stage” is a term to be used for visual information processing and “area” is an anatomically and/or neurophysiologically defined region in the brain. However, here we roughly regard V1 (the first visual cortex to which visual signals from the eyes are projected), V2 and V3 as being in the early stage of visual information processing. These areas are generally called the early visual cortex and known to process more primitive visual signals more locally with smaller receptive fields than in higher visual areas (Hubel & Wiesel 1968, Roe & Ts'o 1995). We regard V4, middle-temporal area (MT or V5) and V3A particularly for humans (Koyama et al 2005, Tootell et al 1997) as being in the mid stage. These areas respond to visual signals more globally than in the early visual cortex. It has been suggested that V4 correlates with color constancy (Zeki 1993) and a surface representation (Bouvier et al 2008). MT (V5) of moneys/humans and V3A of humans have been found to respond to global motion (Braddick et al 2001, Koyama et al 2005, Movshon et al 1985, Newsome & Pare 1988, Rees et al 2000). We regard any cortical areas that process visual signals beyond the mid stage as being the late stage. While the early and mid stages consists of the visual cortex in which visual signals from the eyes are processed and integrated, visual information beyond the visual cortex is processed in more cognitive fashions. The areas include the lateral intraparietal sulcus (LIP) and dorsolateral prefrontal cortex (DLPFC). It has been found that LIP (Shadlen & Newsome 1996) and DLPFC (Heekeren et al 2004) correlate with visual decision.
Early-stage model
What is changed?
This model assumes that task-relevant VPL is associated with changes in the early visual cortex (Adab & Vogels 2011, Adini et al 2002, Bejjanki et al 2011, Choi & Watanabe 2012, Crist et al 1997, Dill & Fahle 1997, Fahle 2002, Fahle & Edelman 1993, Fahle & Poggio 2002, Harris & Fahle 1995, Karni & Sagi 1991, Karni & Sagi 1993, Schwartz et al 2002, Xu et al 2010, Xu et al 2012a, Xu et al 2012b, Xu et al 2012c). A number of studies have shown that VPL is highly specific for the trained location, feature, and eye. With regards to location specificity, psychophysical studies have found that performance improvement on the trained feature that was presented in a specific location of the visual field during training is not observed any more if the same feature is presented even 1 or 2° shifted from the trained location (Ahissar & Hochstein 1997, Crist et al 1997, Fahle & Edelman 1993, Fiorentini & Berardi 1980, Karni & Sagi 1991, McKee & Westheimer 1978, Poggio et al 1992, Saarinen & Levi 1995, Shiu & Pashler 1992, Watanabe et al 2002). Feature specificity has also been found for visual features such as orientation (Fiorentini & Berardi 1980, Poggio et al 1992, Schoups et al 1995), motion-direction (Ball & Sekuler 1987, Koyama et al 2004, Vaina et al 1998, Watanabe et al 2002). Eye specificity indicates that VPL trained with one eye does not completely transfer to the eye in many cases (Fahle & Poggio 2002, Karni & Sagi 1991, Karni & Sagi 1993), although that is not always the case (Schoups & Orban 1996). Since signals from the two different eyes are not converged until V1, incomplete transfer of VPL suggests that VPL is at least partially associated with changes before the conversion that starts in V1, although another interpretation has also been proposed (Young et al 2004).
Based on such specificity it has been suggested that VPL occurs in the early visual cortex, which was once thought to become hard-wired early in the human development process (i.e., so rigid that hardly any change occurred after an early critical period in life). It has been shown that this high specificity of a feature in VPL is in accord with neurophysiological findings that neurons in the early visual cortex tend to possess smaller receptive fields than those in higher areas (Felleman & Van Essen 1991, Zeki 1993). Orientation specificity tends to be higher in cells in low-level visual processing areas (Felleman & Van Essen 1991, Zeki 1993). In addition, it has been reported that external feedback regarding the correctness of a subject's responses (response feedback) is not necessary for VPL to occur (Fahle et al 1995, Poggio et al 1992). The featural specificity and lack of need for feedback led some researchers to suggest that VPL occurs in the early visual system.
Results of some animal physiological studies suggest that changes occur in V1 in association with task-relevant VPL. As a result of training on an orientation discrimination task, responses of neurons most sensitive to differences in the discriminated orientations in monkey V1 increased (Schoups et al 2001). In another study, although neither basic receptive field properties nor visual topography was changed in association with VPL, the influence of contextual stimuli placed outside the receptive field showed a change consistent with results of the trained discrimination (Crist et al 2001). Perceptual learning improved contrast sensitivity in V1 neurons in cats (Hua et al 2010). By means of optogenetics, a cutting-edge technique, in which specific neurons are sensitized by light, activation of a specific subtype of cortical inhibitory interneurons was induced. This activation led the mouse V1 to sharpen neuronal feature selectivity in V1 and improves perceptual discrimination (Lee et al 2012).
A number of studies, including training on a contrast discrimination task (Furmanski et al 2004) and a texture discrimination task (TDT) (Schwartz et al 2002, Walker et al 2005, Yotsumoto et al 2008), have found that VPL is associated with increases in blood oxygenation level dependent (BOLD) signal in the region of V1 in humans that corresponds to the location of the trained stimulus. It has been found that while within the first few weeks of training, performance on TDT increased, BOLD signal in the trained region of V1 increased (Yotsumoto et al 2008). However, after the performance increase saturated, brain activation in V1 decreased to the level observed before training while the high performance was retained. In that case, no systematic activity change particularly in the region or hemisphere corresponding to the trained location was observed in any other brain area. This rules out the possibility that top-down influence or feedback from higher cortical area(s) modulate V1 activation changes. Another study found that during non-rapid eye movement (N-REM) sleep following training of the texture discrimination task, significantly higher BOLD signal was observed in the trained visual quadrant of V1 than in another quadrant (Yotsumoto et al 2009). It has been found that BOLD signals are better correlated with field potential than action potential and reflects the input and intracortical processing of a given area rather than its action potential output (Logothetis et al 2001). This finding might lead one to think that activation in V1, for example, reflects input of activity to V1 from a higher area. However, most research that showed V1 changes did not find any systematic activity change in higher areas. Thus, it is reasonable to assume that BOLD changes in V1 correspond to those in neural circuits within V1.
How is the early stage changed?
It is generally assumed that attention or feedback to V1 or the early visual cortex during training plays an important role in forming task-relevant VPL. Several studies have found cases in which a task-irrelevant feature that is presented while a subject is performing a task on another feature during training does not lead to learning of the task irrelevant feature (Ahissar & Hochstein 1993, Schoups et al 2001, Shiu & Pashler 1992). It has also been found that in some cases perceptual learning of a task on a feature does not transfer to another task on the trained feature (Huang et al 2007, Westheimer et al 2001). These findings suggest that attention/feedback signals play an important role in sending signals that are highly specific for the trained feature and/or task possibly to lower level stages. Ahissar and Hochstein (1993) have found that as task difficulty increases, VPL becomes more specific with respect to both orientation and position and suggested that as perceptual learning proceeds, the stage/cortical area involved in VPL becomes lower. This so-called reversed hierarchical model has been influential in VPL research and other fields in visual sciences (Ahissar & Hochstein 1997, Hochstein & Ahissar 2002). For example, discrimination between two relatively distant orientations can be successfully performed in a high visual stage that process relatively coarse orientation and location signals. However, if fine-grained orientation discrimination is required, a lower visual area that processes finer orientation and location signals needs to be used and learned. To do so, attention is regarded as an important driving factor (Hochstein & Ahissar 2002). Fahle (2009) has also suggested that for easy tasks, elaboration of top-down influences are usually not required, and learning is restricted to late-stage selection of the appropriate signals on higher cortical levels, which seems easier and faster to achieve. However, to reach the absolute limits of sensory performance, VPL seems to optimize the entire chain of sensory processing (Fahle 2009). These models assume that demands to improve a task ultimately require changes in V1 that has high specificity in location and primitive visual features including orientation and spatial frequency. On the other hand, horizontal connections have also been observed within V1 (Ts'o & Gilbert 1988). Gilbert and his colleagues (Piech et al 2013) assume that feedback signals driven by a task demand interacts horizontal connections in V1 and change the local gains in V1 which are associated with perceptual learning. Note that although these models assume an important role of attention in task-relevant VPL, they do not assume that the attentional system or higher-level cognitive processing related to attention is changed in association with task-relevant VPL.
Mid-stage model
What is changed?
Some studies have suggested that VPL is not associated with changes at the early stage including V1, but with changes at the mid stage. As aforementioned, the mid stage includes middle-temporal area (MT), V4 and V3A particularly for humans (Koyama et al 2005, Tootell et al 1997). Here we will review psychophysical and neuroscientific (animal studies and human brain imaging) studies.
In psychophysics, it has been found that some types of VPL depend on perceptual constancy that corresponds to stable representation of certain properties of an object, despite variable visual input. Such representation is thought to occur in the middle visual stage (Garrigan & Kellman 2008). Although training on a task of discriminating close motion directions or orientations leads to VPL specific to the trained direction (Ball & Sekuler 1987) or orientation, learning by training in discriminating larger direction or orientation differences transfers to untrained directions (Liu 1999). These results suggest that in some conditions VPL occurs in the mid stage in which location and feature signals are less specific than V1. As aforementioned, the reversed hierarchy model (Ahissar & Hochstein 1997, Hochstein & Ahissar 2002) indicates that VPL proceeds from higher to lower stages including V1. It would be natural to assume that VPL starts with some middle level stage. Note that in psychophysics the distinction between the mid and late stages is not highly clear and results that seem to support the mid stage changes could also be interpreted as changes in the late stage.
In animal studies, in V4 of monkeys, tuning properties changed in association with VPL of an orientation discrimination task (Yang & Maunsell 2004). However, no tuning change was observed in V1 (Ghose et al 2002, Yang & Maunsell 2004). In another monkey study (Chowdhury & DeAngelis 2008), inactivation of MT by injection of the GABA agonist muscimol prior to coarse absolute depth discrimination training was shown to impair performance. However, fine relative depth discrimination training with MT inactivation neither impaired coarse depth discrimination nor led to changes in the disparity tuning of MT neurons. The authors concluded that when MT activation is inhibited, other areas in the ventral visual pathway are recruited and mediate coarse depth discrimination.
It has been found that VPL of discrimination of coherent motion directions is associated with changes in V3A of humans. Behavioral tuning function changes after global motion detection training were compared with decoded tuning function changes for 8 visual areas using pattern classification analysis BOLD signals (Kamitani & Tong 2005). The behavioral tuning function changes were extremely highly correlated to decoded tuning function changes only in V3A, which is known to be highly responsive to global motion with human subjects (Braddick et al 2001, Koyama et al 2005, Shibata et al 2012a).
How is the mid stage changed?
Dosher and Lu have built a reweighting model of perceptual learning, which has been highly influential in VPL research (Dosher & Lu 1998, Liu et al 2010, Lu et al 2005, Petrov et al 2005, Petrov et al 2006). The model assumes that VPL occurs due to two independent mechanisms, improved filtering of external noise and removal of internal noise. They assumed that selective re-weighting or “readout” of connections between V1 and a decision unit (that is, the mid stage) leads to simultaneous improvements of these two noises (Dosher & Lu 1998). V1 was regarded as a place for stimulus representation in which spatio-temporal filtering occurs. This model assumes no change in the stimulus representation and therefore does not support the model in which task-relevant VPL occurs due to changes in the stimulus representation and therefore V1 (Petrov et al 2005). Recently, the model has been modified to have two levels of representations, location specific and location-independent representations (Dosher et al 2013) to explain location transfer demonstrated by double training (Wang et al 2012, Xiao et al 2008, Zhang et al 2010a) which is discussed in the late-stage section. Based on the analysis of data indicating that responses of neurons in LIP, but not in MT, were changed in association with VPL of coherent motion (Law & Gold 2008), the Gold group has suggested weighting changes occur between MT and LIP (Gold et al 2008). While these reweighting models do not assume changes in V1, it has been pointed out that theoretically speaking, improvement in the efficiency of probabilistic inference in early cortical circuits by reweighting connections within the early visual cortex can explain noise reduction and tuning curves changes in V1 and V4 (Bejjanki et al 2011).
Late-stage model
Based on a number of recent physiological and psychophysical studies, the role of the late stage, which is defined as all processing higher than mid-stage, in task-relevant VPL has been much more emphasized.
What is changed?
As a result of training on a shape-identification task, resting state functional connectivity and directed mutual interactions between trained mid-visual areas and frontal-parietal areas involved in the control of spatial attention were modified (Lewis et al 2009). In addition, in association with VPL of a visual discrimination task, individual differences in performance changes due to the training are related to spontaneous cortical activity between early- and mid-level visual cortices and also between visual cortex and prefrontal association areas. These results suggest that changes associated with task-relevant VPL include changes in connectivity involving an attentional source system (Baldassarre et al 2012). Another study found that behavioral improvements in an orientation discrimination task occur with activity changes in the Anterior Cingulate cortex (ACC), suggesting that perceptual learning is associated with a high-level cognitive area (Kahnt et al 2011).
Gold and his colleagues found that perceptual learning of coarse coherent motion direction discrimination was associated with tuning property changes in LIP, an area known to be involved in perceptual decision making (Shadlen & Newsome 2001), but not in MT, an area belonging to the mid stage (Law & Gold 2008). As discussed in the mid stage model section, the model proposed by this group assume reweighting in connections between MT and LIP (Gold et al 2009). However, another possibility that is not mutually exclusive to the reweighting model is that at least VPL of coarse motion direction discrimination is associated with changes in decision factors including a decision criterion, which is reflected in LIP response changes (Shadlen 2014).
In psychophysics, there has been a breakthrough finding with regards to transfer of VPL. Training on a feature (e.g., contrast) at one location and additional training with an irrelevant feature/task (e.g., orientation) at a second location resulted in a complete transfer of learning of the feature (e.g., contrast) to the second location (Xiao et al 2008). Based on a series of interesting experiments by the group led by Cong Yu (Wang et al 2012, Xiao et al 2008, Zhang et al 2010a), this research group has built a model in which task-relevant VPL is associated only with changes in the late stage that is involved in the source of attention from which attentional signal is exerted and decision making. In this model, location specificity of task-relevant VPL results from long-term inhibition from the cognitive system on all locations in the visual field except the trained location. When training on a second task occurs in a different location than the previously trained location, the connection between the late stage and this location is reactivated and the inhibition to the location is released. As a result, VPL of the original task was transferred to the location that was trained for the second task (Xiao et al 2008). As discussed in the early-stage model, the model by the Sagi group also suggests that transfer of task-relevant VPL results from the abolishment of the link between the early stage and late stage (Harris et al 2012). However, the models by the Yu group and the Sagi group are fundamentally different. While the Yu group assumes that the link between the early and late stages has been made and kept by the late stage (Xiao et al 2008), the Sagi group assumes that by the link the location signals from the low stage constrain location specificity of task-relevant VPL (Harris et al 2012).
Summary of Task-Relevant Visual Perceptual Learning
Models of task-relevant VPL are roughly classified into 3 different types, which we call the early-stage model, mid-stage model, and late-stage model, according to the stage at which processing or mechanisms is changed in association with task-relevant VPL.
One possible reason that there are such a variety of models is that many different studies have used highly different experimental conditions with different parameter sets. Despite the variety of methods employed in this body of research, many studies overgeneralize their results and draw conclusions as if their specific results can be applied to all types of VPL. However, it is not either certain or even likely that all types of task-relevant VPL are subserved by only one stage. The variance of experimental parameters leaves the possibility that VPL is subserved by multiple mechanisms.
Task-Irrelevant Visual Perceptual Learning
What is Task-Irrelevant Visual Perceptual Learning?
The previous section focuses on task-relevant VPL in which performance increase on a task and/or feature occurs as a result of training on the task and/or the feature. In 2001 it was found that performance increase also occurs as a result of mere exposure of a feature that is irrelevant and subthreshold to a given task during training (Watanabe et al 2001). In that study, 5% coherent motion was presented in the background while subjects repeatedly performed an rapid serial visual presentation (RSVP) task that required them to report 2 white letters in a sequence of otherwise black letters. In the pre- and post tests, the performance on the same exposed 5% coherent motion direction were around chance level and therefore subthreshold, whereas a significant performance improvement was observed on the same direction with 10% coherency that is above-threshold. These results indicate that mere exposure to coherent motion that is task-irrelevant and subthreshold induces learning. It is difficult for focused attention to be kept on such a task-irrelevant and subthreshold direction. Therefore it was concluded that perceptual learning of a feature occurs without enduring focused attention being directed to the feature. We call this new type of VPL task-irrelevant VPL. A number of subsequent studies have reported task-irrelevant VPL (Barbot et al 2011, Baumann et al 2008, Beste & Dinse 2013, Beste et al 2011, Carrasco et al 2008, Gutnisky et al 2009, Leclercq et al 2013, Leclercq & Seitz 2012, Rosenthal & Humphreys 2010, Seitz & Watanabe 2005, Seitz & Watanabe 2003, Seitz et al 2005b, Tsushima et al 2008, Watanabe et al 2002, Xu et al 2012b, Zhang & Kourtzi 2010, Zhang et al 2010b).
Reinforcement and Task-Irrelevant Visual Perceptual Learning
It has been later found that task-irrelevant VPL at least in the similar setting as in the 2001 study by the Watanabe group does not occur without reinforcing factors involved. Task-irrelevant VPL occurs only when the exposed 5% coherent motion direction was paired with a target in RSVP task (white letter) (Seitz & Watanabe 2003) and only when subjects made correct detections on targets, which might cause internal reward or satisfaction (Seitz et al 2005a). Furthermore, VPL of the task-irrelevant 5% coherent motion direction occurred when the feature was paired with reward and did not occur without reward (Seitz et al 2009).
Based on these findings, Seitz and Watanabe built a unified model, in which task-relevant VPL as well as task-irrelevant VPL results from interactions between spatially diffusive reinforcement signals and bottom-up signals from an exposed stimulus feature (Fig. 1). Reinforcement signals arise when subjects are given external reward or when they have conducted successful task performance. The bottom-up signals are either from a task-relevant or task-irrelevant feature. In that schedule, task-relevant and task-irrelevant VPL share the same underlying principles (Seitz & Watanabe 2005).
Figure 1.

The Seitz & Watanabe model of VPL. The model assumes that VPL results from interactions between spatially diffusive reinforcement signals that are triggered by successful task performance and bottom-up signals from a feature presented during training, irrespective of whether the feature is task-relevant or task-irrelevant. Attention enhances bottom-up signals from a task-relevant feature, whereas it decreases or suppresses signals from a task-irrelevant feature. A red arrow represents excitatory stimulation and a blue arrow inhibitory stimulation. Task-irrelevant VPL occurs only when task-irrelevant feature signals are so weak that the signals fail to be detected and therefore are not suppressed by an attentional system. However, it is noteworthy that if a task-irrelevant feature is presented in an environment optimal to the feature, task-irrelevant learning can occur if the feature is supra-threshold. Task-relevant VPL and task-irrelevant VPL are location specific due to constraints from the bottom-up signals from a feature presented during training.
Attention and Task-Irrelevant Visual Perceptual Learning
However, later findings have suggested that the mechanism or processing which leads to task-irrelevant VPL is not exactly the same as task-relevant VPL. There are several aspects that could reveal differences between mechanisms of the two types of VPL. First, there are differential effects of cognitive control on the two types. In one study, the coherence (and therefore detectability) of a task-irrelevant motion signal was varied during the performance of a central task (Tsushima et al 2008). Task-irrelevant VPL occurred only when the coherent motion signal was below or close to the detection threshold. This result is consistent with a model in which a cognitive control system fails to detect weak or below-threshold task-irrelevant signals, and therefore fails to suppress these signals, thus allowing them to exert a stronger influence on information processing than stronger task-irrelevant signals that can be detected by the attentional system (Tsushima et al 2006, Tsushima & Watanabe 2009).
At the same time, it is noticeable that in conditions in which task-irrelevant features are not suppressed by the attentional system, task-irrelevant learning occurs. For example, if a task-irrelevant feature is presented spatially distant from a task-relevant feature, learning occurs on the task-irrelevant feature (Watanabe et al 2002). In a configuration in which the same two features, one task-relevant and the other task-irrelevant, are presented at different locations, feature-based attention could enhance both features so that task-irrelevant learning may occur (Gutnisky et al 2009). Exposure to passive visual low-frequency supra-threshold stimulation of irrelevant orientation improved subsequence performance on the orientation (Beste et al 2011). Seitz and Dinse have suggested that task-irrelevant features could occur in a condition in which a stimulus environment is optimal to task-irrelevant feature if it is not weak or subthreshold (Seitz & Dinse 2007, Zhang et al 2010b) and that the optimal environments to task-irrelevant features are not the same as task-relevant features.
Second, task-involvement of the trained feature in task-relevant VPL makes its mechanism distinguished from task-irrelevant VPL. If task-relevant VPL is at least partially associated with changes in processing of the trained task, the mechanisms of task-relevant VPL should not be the same as the mechanisms of task-irrelevant VPL. As discussed in the previous section, in some cases VPL of one task on a feature does not completely transfer to a different task on the same or similar feature (Huang et al 2007, Westheimer et al 2001). This indicates that task-relevant VPL is at least partially associated with changes in the mechanism specific for the trained task, which should not be shared with task-irrelevant VPL.
In another study, double dissociation between task-relevant and task-irrelevant VPL was observed (Beste et al 2011). First, subjects were asked to conduct a change-detection task on luminance changes against orientation changes as task-irrelevant. This task performance was followed by exposure to passive visual high- or low-frequency stimulation of either the relevant luminance or irrelevant orientation feature. High-frequency stimulation using luminance enhanced performance to detect luminance changes, whereas low-frequency stimulation impaired performance. In contrast, while low-frequency exposure of orientation as task-irrelevant impaired performance, high frequency orientation stimulation enhanced performance. These results also suggest task-irrelevant VPL does not share exactly the same mechanism as task-relevant VPL.
Using a new, functional magnetic resonance imaging (fMRI) online-feedback method, patterns of activation in early visual areas corresponding to the pattern evoked by the presentation of a real and specific target orientation stimulus were induced in early visual cortex (Shibata et al 2011). These patterns were repeatedly induced without external stimulus presentation and without subjects' knowledge of what was being induced. As a result, a significant sensitivity only to the target orientation improved. That is, mere repetition of an activity pattern in the early visual cortex corresponding to a feature may be sufficient to induce VPL of that feature. The results also indicate that at least the plasticity with no direct involvement of task training can occur in the early stage. This plasticity could manifest as task-irrelevant VPL.
There are studies that compared activations that are related to task-relevant and task-irrelevant learning. One study measured BOLD signals evoked when subjects discriminated the spatial frequency of gratings presented with the same or orthogonal orientation (Baumann et al 2008). The BOLD signals were significantly higher in the early visual areas such as V1 and V2 when the gratings to be compared had an orthogonal orientation, compared to when they had the same orientation. In contrast, the task-irrelevant stimulus property (orientation) had no significant effect on the prefrontal and intraparietal cortex. Combining the results of these studies, it may be plausible to think that task-irrelevant VPL is associated with changes in early stage, whereas learning that results from involvement of a task results is associated with changes in higher stages.
Summary of Task-Irrelevant Visual Perceptual Learning
Task-irrelevant VPL has been found more recently than task-relevant VPL. It has been suggested that both task-relevant and task-irrelevant VPL are at least partially result from interactions between reinforcement signals and bottom-up signals from the presented feature irrespective of whether the feature is task-relevant or task-irrelevant (Seitz & Watanabe 2005). At the same time, some different aspects of mechanisms seem to be involved in these two types of VPL. One is due to differential effects of cognitive control. A task-relevant feature is enhanced by attention while task-irrelevant features may be suppressed (Tsushima et al 2006, Tsushima et al 2008, Tsushima & Watanabe 2009). Thus, unless a given environment is optimal to task-irrelevant VPL, task-irrelevant VPL does not occur if it is subthreshold and therefore cannot be detected by the source of attention. Another aspect is that while task-relevant VPL may be at least partially associated with changes in processing specific for the trained task, task-irrelevant VPL is not. Results of recent human imaging studies (Baumann et al 2008, Shibata et al 2011, Zhang et al 2010b) suggest task-irrelevant VPL is associated with the early stage while changes resulting from training on a task are associated with higher stages. Brain imaging research results suggest that task-irrelevant VPL may be associated with changes in the low stage.
Dual-Plasticity Model of Visual Perceptual Learning
Background
While a vast number of results concerning VPL have accumulated, there is no consensus about the mechanism. An unfortunate tendency is that advocates of one model of task-relevant VPL deny the evidence of one model over the other and build a detailed model supporting only one view of the aforementioned controversies. In addition, none of the models of task-relevant VPL can satisfactorily explain task-irrelevant VPL. Since it is highly unlikely the basic mechanisms of task-relevant and task-irrelevant VPL are completely different, it is necessary to build a model that can explain both task-relevant and task-irrelevant VPL.
To resolve these issues, here we propose a dual plasticity model of VPL which could resolve the issues and account for most of the accumulated data that include those appearing to contradict each other. The dual plasticity model will incorporate the controversy as to what stage is associated with task-relevant VPL and the mechanisms of both task-relevant VPL and task-irrelevant VPL. First we will describe the dual plasticity model, and then discuss the rationalizations for the framework.
Assumptions of dual plasticity model
Here we propose that there are at least two different types of plasticity, feature-based plasticity and task-based plasticity, which result from different processings during training, include changes in different stages, and occur at different timings (Fig. 2). Feature-based plasticity is defined as changes in a representation of features. Task-based plasticity is defined as changes in processing related to a trained task.
Figure 2.

Schematic illustration of the dual plasticity model. In this model, there are two types of plasticity, feature-based plasticity and task-based plasticity. Feature-based plasticity results from presentation of a feature during training, irrespective of whether the feature is task-relevant or task-irrelevant. Feature-based plasticity reflects changes in feature representation and constrains VPL to the trained feature and location. Task-based plasticity arises as a result of involvement of a given task during training. Task-based plasticity reflects changes in task processing and constrains VPL to the trained task. Task-relevant VPL (R-VPL) consists of feature-based plasticity and task-based plasticity. Task-irrelevant VPL (I-VPL) consists of only feature-based plasticity.
How do feature-based plasticity occur?
We assume that how feature-based plasticity occurs follows our reinforcement model (Seitz & Watanabe 2005). In this model, both task-relevant and task-irrelevant features are learned as a result of interactions between spatially diffusive reinforcement signals and bottom-up signals from a feature exposed during training, irrespective of whether the feature is task-relevant or task-irrelevant (Fig. 1). Basic changes in representation of the exposed feature are constant, irrespective of whether the feature is task-relevant or task-irrelevant and whether of what kind of task on the feature is trained if the feature is task-relevant. However, the overall amplitude of changes including no change depends on the rules of cognitive control, as discussed in the low-stage change section. If an exposed task-irrelevant feature is strong, the feature is detected and suppressed by a cognitive control system if a stimulus environment is optimal to the feature. If the feature is weak and is not detected by the cognitive control system, the feature is not suppressed and feature-based plasticity on the feature occurs (Tsushima et al 2006, Tsushima et al 2008). On the other hand, a relevant-feature is enhanced rather than being suppressed by the cognitive control system. While Seitz and Watanabe (2005) assumed that the reinforcement model are for the whole model of task-relevant and task-irrelevant VPL, here we propose that this reinforcement mechanism is for the feature-based plasticity (Fig. 3). To explain the whole mechanism of VPL, we now believe that not only feature-based plasticity but also task-based plasticity and its interactions with feature-based plasticity are necessary to be assumed.
Figure 3.

Model of the mechanism of feature-based plasticity. While the Seitz & Watanabe model assumes that this is the entire mechanism of both task-relevant and task-irrelevant VPL, here we propose that the mechanism of the Seitz & Watanabe model is only for feature-based plasticity. Task-relevant VPL is subserved by this feature-based plasticity mechanism and also by task-based plasticity that results from performing a given task during training and is also specific for the trained task. A red arrow represents excitatory stimulation and a blue arrow inhibitory stimulation.
How does task-based plasticity occur?
Task-based plasticity occurs as a result of involvement of a trained task rather than exposure to a feature as in the feature-based plasticity. Thus, training on different tasks (say, detection task vs discrimination task) with the same feature (say, orientation) can lead to qualitatively different changes in task-based plasticity.
Where do feature-based plasticity and task-based plasticity occur?
We assume that feature-based plasticity is a change in a feature representation and occurs in the early or mid stages. Whether it occurs at the low or mid stage depends on the feature on which feature-based plasticity occurs. If a feature is highly primitive and a representation of the feature is first built in V1, feature-plasticity of the feature should occur in V1 that belongs to the low stage. If a feature is made of components of more primitive features, feature-based plasticity of the feature may occur in the mid stge. For example, orientation starts being processed in V1 in the cortex (Hubel & Wiesel 1968). Thus, the areas in which feature-based plasticity occurs may include V1. Coherent motion consists of coherent moving dots and randomly moving dots from one temporal frame to another. Coherent motion starts being processed distinctively from random motion in V3A/MT in humans (Braddick et al 2001, Koyama et al 2005, Rees et al 2000) and MT in monkeys (Newsome & Pare 1988). Thus, the areas in which feature-based plasticity of coherent motion occurs may include V3A/MT in humans and MT in monkeys.
Feature-based and task-based plasticity and task-relevant and task-irrelevant VPL
We assume that task-relevant VPL consists of both feature-based plasticity and task-based plasticity, whereas task-irrelevant VPL consists of only with feature-based plasticity. Thus, task-relevant and task-irrelevant VPL partially shares the same mechanism, that is, the mechanism of feature-based plasticity.
Specificity and transfer of feature-based plasticity and task-based plasticity
Feature-based plasticity is specific for an exposed feature and the location in which the feature was presented during training. Task-based plasticity is specific for the trained task. Task-irrelevant VPL is constrained by the specificity from the feature-based plasticity (feature and location). Task-relevant VPL is constrained by the specificity from the feature-based plasticity (feature and location) and also by the specificity from the task-based plasticity. If feature-based specificity is abolished in task-relevant VPL as a result of an experimental manipulation, the feature-based constraints should be eliminated. As a result, VPL of the trained task should be transferred to different locations/features (Harris et al 2012). If task-based plasticity is eliminated in task-relevant VPL of a feature, VPL of the feature should be transferred to other tasks.
Empirical Studies for Dual Plasticity Model
There are several significant studies whose results are in accord with the dual plasticity model. First, in one study (Watanabe et al 2002), during training an RSVP task was performed at the center of the display while a global motion stimulus, in which local dots moved in random directions within a certain range, was being presented in the background (Watamaniuk et al 1989). There are two types of perceived motion in this type of stimulus: the local motion signals of the individual dots and a global motion direction whose direction corresponds to the spatio-temporal average of local motion signals in the display. The range of local motion directions was varied across different subject groups. As a result, performance increased for all measured motion directions within the range of local motion, but not for the global motion percept. This indicates that VPL occurred for task-irrelevant local motion directions, but not for the global motion direction. In another experiment of the same study, during training, a discrimination task on global motion direction was conducted. In this condition, the global motion direction was task-relevant. In the earlier phase of training, performance increased within the range of local motion, as in the first experiment in which global motion was task-irrelevant. However, in the later phase, performance improved only around the trained directions. From these results, it was concluded, “The learning gain for a range of local motion was obtained weather global motion was task-relevant or task-irrelevant. In a later phase, there is learning gain only for a task-relevant global motion direction; that is, only in response to task demands (task-irrelevant VPL of exposed local directions) may occur only at a very low-level stage and seems to be processed independently of global motion, whether global motion is task-relevant or irrelevant. In contrast, task-relevant VPL may occur at multiple stages (p 5 in Watanabe et al, 2002).” These results are in accord with the dual plasticity model. That is, the learning gain for a range of local motion is regarded as manifestation of feature-based plasticity and learning gain only for a task-relevant global motion direction as a manifestation of task-based plasticity.
Second, the Sagi group applied an operation that abolishes orientation adaptation to the orientation trained by the TDT and found that the location specificity observed in VPL of TDT was eliminated (Harris et al 2012). This finding is in accord with the dual plasticity model. The results suggest that a type of learning in the early stage constrains the location of VPL to the location of the trained stimulus, whereas another type of learning is related to improvement in task performance (Harris et al 2012). These proposed two types of learning can correspond to feature-based and task-based plasticity of task-relevant VPL.
Third, learning of a collinearity (regularities in natural contours) occurs by simple exposure without task involvement and is associated with activation by occipitotemporal areas (low and mid stages) whereas learning to integrate line segments orthogonal to contour paths requires specific training and also is associated with activity enhancement in intraparietal regions that mediate attentional processing (Zhang et al 2010b). Although this study used different stimuli in different conditions and cannot rule out the possibility that the line segments orthogonal to contour paths themselves are processed in higher stages than collinear segments, the results suggest that learning due to training with a task is associated with higher stages than training without the task.
Recently Shibata and his colleagues have conducted a brain imaging experiment to test the validity of an important aspect of the dual plasticity model (Shibata et al 2013). In test stages that were conducted before and after the training on detection of coherent motion, BOLD signals were measured in two conditions: a feature-based condition and a task-based condition. In the feature-based condition, a central RSVP task was conducted while the same motion as the trained were presented as task-irrelevant. In the task-based condition, the same task on the trained motion as in training was conducted. The results showed that V3A that is known to be highly responsive to coherent motion (Braddick et al 2001, Koyama et al 2005) was significantly more activated in the post-test than in the pre-test in both the feature-based and task-based conditions. On the other hand, multiple areas including the intraparietal sulcus, which includes the human homologue of monkey LIP, were significantly more activated in the post-test than in the pre-test only in the task-based condition. These results are in accord with a key aspect of the dual plasticity model. That is, feature-based plasticity is associated with the changes in representation of a feature, which should be manifested when the feature is presented irrespectively of whether the feature is task-relevant or task-irrelevant. Task-based plasticity is associated with the changes in processing related to the trained task, which should be manifested only while the same task as in training is conducted.
Dual Plasticity Model and Existing Models
How does task-relevant VPL occur?
The dual plasticity model is distinguished from other models. With regards to the question of how task-relevant VPL occurs, there are at least two aspects in the dual plasticity model that are clearly distinguished from other models of VPL. First, no other model is as comprehensive as to explain both task-relevant VPL and task-irrelevant VPL. In the dual plasticity model, task-relevant VPL is associated with both feature-based plasticity and task-based plasticity, whereas task-irrelevant VPL is associated only with feature-based plasticity. Second, to our knowledge all other models assume that task-relevant VPL results from task-involvement. These models do not consider the possibility that even task-relevant VPL results from exposure to the trained feature as in the dual plasticity model.
What is changed?
With regards to what is changed, most models assume that task-relevant VPL is associated with changes at only one of the early, mid and late stages. There is a general tendency that results that support models assuming changes in one stage are ignored by researchers who support models assuming changes in another stage. In contrast, the dual plasticity model assumes that task-relevant VPL is associated with changes in more than one stage, which could comprehensively explain apparently conflicting results.
As discussed above, many early stage models assume that what is changed is a representation of the trained feature, based on experimental results that have shown high specificity of VPL for the trained location, feature, and eye, as well as results indicating that VPL is associated with changes in responses in early visual areas (Adini et al 2002, Crist et al 1997, Dill & Fahle 1997, Fahle 2002, Fahle & Edelman 1993, Fahle & Poggio 2002, Gilbert 1994, Harris & Fahle 1995, Karni & Sagi 1991, Karni & Sagi 1993, Xu et al 2010, Xu et al 2012a, Xu et al 2012b, Xu et al 2012c, Yang & Maunsell 2004) (but see (Bejjanki et al 2011)). Double training leads to transfer of one type of task-relevant VPL to another location at which another task is trained or a feature is exposed. It is difficult for these early stage models to explain transfer of location and features (Wang et al 2012, Xiao et al 2008, Zhang et al 2008, Zhang et al 2010a, Zhang et al 2010b). In addition, it is impossible for the early stage models alone to explain response changes in higher areas (Baldassarre et al 2012, Kahnt et al 2011, Law & Gold 2008).
Most mid-stage and late-stage models assume that what is changed is weighting connecting different areas (Chowdhury & DeAngelis 2008, Dosher et al 2004, Dosher & Lu 1998, Dosher & Lu 1999, Gold et al 2009, Gu et al 2012, Kahnt et al 2011, Lewis et al 2009, Liu et al 2010, Petrov et al 2005, Xiao et al 2008, Zhang et al 2008, Zhang et al 2010a, Zhang et al 2010b). However, these models on their own cannot explain the results that V1 is changed in association with task-relevant VPL or incomplete inter-ocular transfer (transfer of VPL trained with one eye to the other eye) (Karni & Sagi 1991, Karni & Sagi 1993, Shibata et al 2012b, Xu et al 2012b).
The dual plasticity model can comprehensively explain changes at the low-, mid- and late stages.
Early-stage models and dual plasticity model
Most results that have been regarded as evidence for the early-stage model are in accord with the feature-based plasticity of our dual plasticity model, which corresponds to changes in a representation of a feature presented during training. Specificity of the trained feature including its location is in accord with the hypothesis that such specificity is in the changed representation and constrains the performance of VPL (Harris et al 2012). Tuning curve changes in the early and mid stage areas (Hua et al 2010, Schoups et al 2001, Yang & Maunsell 2004) may be related to changes in the representation. BOLD signal enhancement in the trained location (Furmanski et al 2004, Schwartz et al 2002, Walker et al 2005, Yotsumoto et al 2008) is also consistent with the hypothesis that feature-based plasticity occurs in an earlier-stage than task-based plasticity. In addition, an on-line fMRI neurofeedback method repetitively induced the activations only in the subjects' early visual cortex that corresponded to the pattern of activation evoked by the presentation of a real target orientation. As a result, VPL of the orientation occurred. This finding is in accord with some properties of feature-based plasticity including changes in the early visual cortex and no task involvement (Shibata et al 2011).
Mid- and late- stage models and dual plasticity model
As discussed above, it is difficult for the low-stage model or reweighting of the mid-stage model to explain transfer from the trained to untrained locations, unless there is an additional stage or processing where no location specificity is assumed (Dosher et al 2013). In the dual plasticity model, transfer of VPL to a new location could be interpreted as a result of development of a new link between the new location and the task-based plasticity. If the link between feature-based plasticity and task-based plasticity is completely eliminated, task-relevant VPL should have no specificity.
One of the most influential models that assume changes in the mid-stage is the re-weighting model (Dosher & Lu 1998, Gold et al 2008, Liu et al 2010, Lu et al 2005, Petrov et al 2005, Petrov et al 2006). The dual plasticity model does not contradict the reweighting principle. Task-based plasticity can occur as a result of changes in task-driven processing that could occur as a result of re-weighting between visual areas and cognitive areas.
Summary of Dual Plasticity Model
The dual plasticity model consists of feature-based plasticity and task-based plasticity. Feature-based plasticity results from changes in a representation of a feature presented during training, irrespective of whether it is task-relevant or task-irrelevant. Task-based plasticity arises as a result of involvement in a task, irrespective of the trained feature. Task-relevant VPL consists of feature-based plasticity and task-based plasticity. In this case, feature-based plasticity constrains the trained location of task-relevant VPL. On the other hand, task-irrelevant VPL consists only of feature-based plasticity. Thus, task-relevant VPL and task-irrelevant VPL are not completely different, but are commonly subserved by feature-based plasticity.
The dual plasticity model is still a schematic model. Detailed mechanisms of the two types of plasticity and links of these two types have yet to be developed. However, currently most mechanistic models are based on one or two of the three views (early-, mid- and late- stage models) and cannot explain empirical results supporting the other view(s). Moreover, none of the mechanistic models of VPL can explain both task-relevant and task-irrelevant VPL. Thus, it would be more fruitful and constructive to build a schematic but comprehensive model first.
Conclusions and Future Research Directions
Researchers hoped that accumulation of VPL data would lead to development of understanding general rules of VPL, which would help to establish a general mechanism of visual plasticity at the neuronal level. Given this hope, a large number of experiments have been conducted. The results have shown a tendency opposite to finding a general rule(s).
One reason is that to date, different researchers have tended to use different sets of parameters as their stimuli, tasks, etc. in conducting their VPL studies. This practice has made it very difficult to make direct comparisons between the findings and to find, if any, general rules of VPL A variety of results have been obtained under various conditions. In some cases contradicting conclusions were drawn from opposite results. If a new method and/or a new stimulus are used and new results of VPL are obtained, in some cases it is difficult to determine whether the new results reveal a hidden important principle/mechanism of VPL or a trivial variation from an established principle due to the use of the new procedure and/or stimulus. For future studies, it is desirable that researchers will use one of a few standard methods as in experimental physics (Klein 2007).
The second reason, which is not unrelated to the first, is that there have not been strong efforts to organize these divergent results toward identification of any possible general rules.
In the future, it is desirable that experiments will be conducted using standard methods to test the validity of aspects of a comprehensive model, rather than emphasizing one narrow view that explains only a portion of accumulated data.
This review primarily focuses on task-relevant VPL and task-irrelevant VPL of primitive features such as motion and orientation. However, research results and framework made from the results would lead to a better understanding of the general mechanisms of task-driven and/or passive learning/memory. For example, passive characteristics of learning as in task-irrelevant VPL have been shown in learning of objects (Hammer et al 2012, Li & DiCarlo 2008, Li & Dicarlo 2012, Witthoft et al 2009), categorization learning (Folstein et al 2010), word learning (Dewald et al 2011, Dewald et al 2013), speech learning (Vlahou et al 2012), general learning (Reed et al 2011). At the same time, VPL of primitive features may have aspects distinguished from these types of memory and learning. Some differences may be due to the fact that early stages of processing primitive visual features are significantly different from other types of learning. Future research should include efforts to find common principles of VPL of primitive features and other types of learning and memory and unique aspects of VPL of primitive features.
Figure 4.
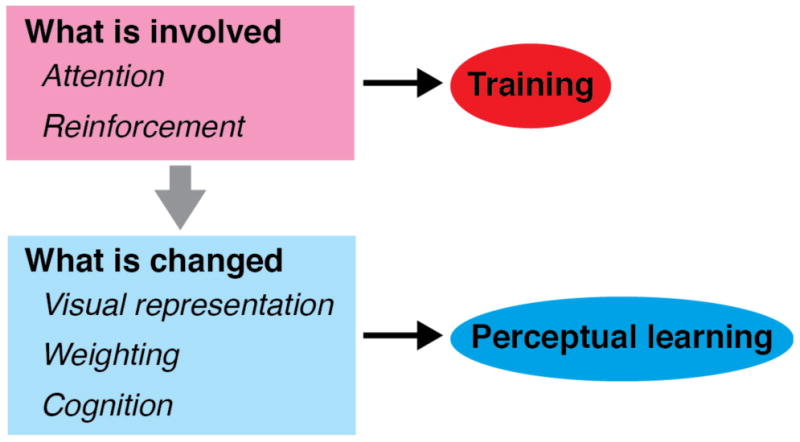
Distinction between what is involved during training on a task and what is changed in association with VPL of the task. What is involved is not necessarily the same as what is changed.
Side Bar 1.
Do response changes in a brain area shown after training always indicate that plasticity has occurred in this area?
Assume that as a result of training on a feature, responses to the feature in a cortical area (say, V1) have changed. In this case, it is premature to conclude that plasticity has occurred in V1. There is at least one more possibility. That is, the changes in V1 could result from temporal feedback from a higher cortical area (say, V4) in which plasticity has actually occurred. In this case, the changes in V1 are not due to plasticity in V1 but merely reflect response changes in V4 due to plasticity. There are at least two ways to rule out this possibility. One is to test whether the response changes in V1 are correlated with response changes in any other area. If there is no correlation between response changes in V1 and any other area, it is likely that the plasticity has occurred in V1. In one study, BOLD signal changes in V1 observed for the first several weeks during training had no correlation with response changes in any other measured area (Yotsumoto et al 2008). The results suggest that the activation changes in V1 are not likely to reflect plasticity in other measured areas. Another way is to examine eye specificity, This may be applied only to the case in which plasticity has occurred in V1. Some types of VPL result from training on a stimulus presented with only one eye (trained eye) and do not transfer to the untrained eye. In such a case, if response changes in V1 are observed when tested with the trained eye while they are not when tested with the untrained eye, this may suggest that plasticity related to the VPL occurred in V1 (Karni & Sagi 1991, Karni & Sagi 1993, Shibata et al 2012b, Xu et al 2012b), although another interpretation has also been suggested (Young et al 2004). This is based on the fact that most monocular cells are found in V1 and feedback from an area higher than V1 does not specifically affect the group of V1 neurons that respond to signals from one eye.
Side Bar 2.
“What is involved?” vs. “What is changed?”
In some cases what is involved during training is confused with what is changed by training (Fig. 4).
During training on a visual task, attention and/or reinforcement processing may be involved in order for subjects to perform the task. For example, if subjects are asked to judge whether two successively presented orientations are the same or different in a task during training, they have to direct attention to the presented orientations. In this case, there is no doubt that focused attention is involved. This indicates that the source mechanism of focused attention is used and areas corresponding to that specific mechanism are activated. However, this does not necessarily indicate that VPL is caused or associated with changes in the attention source mechanism. Likewise, in some cases during training reinforcement processing may play an important role in forming VPL. However, this does not necessarily indicate that reinforcement processing itself is changed in association with VPL.
Summary Points.
VPL is classified into task-relevant and task-irrelevant VPL. Task-relevant VPL of a feature results from training on a given task on the feature, whereas task-irrelevant VPL of a feature arises as a result of exposure to the feature that is irrelevant to a given task.
The stage or brain area that is changed in association with task-relevant VPL is controversial. There are roughly three models, early-stage, mid-stage and late-stage models. A different set of data tends to be used to support each of these models.
Specificity of the trained location and feature was regarded as evidence for the early-stage model. However, it has been recently found that at least in some cases double training results in transfer of VPL to other locations and features. These results suggest that VPL is not associated solely with changes in the early visual cortex or early stage.
A reinforcement model of VPL indicates that both task-relevant and task-irrelevant VPL result from interactions of spatially diffusive reinforcement signals and bottom-up signals from a presented feature, irrespective of whether the feature is task-relevant or task-irrelevant. However, this model alone does not explain either transfer of location/feature after double training or differential results from training on different tasks.
Attention to a task-relevant feature may enhance signals from the feature and increase the probability of the occurrence of task-relevant learning of the feature. On the other hand, if task-irrelevant feature signals are sufficiently strong to be detected by an attentional system, they may be suppressed and cause no task-irrelevant VPL. If task-irrelevant signals are not sufficiently strong to be detected, they may not be suppressed and cause task-irrelevant VPL. At the same time, if a task-irrelevant feature is presented in an environment that optimizes task-irrelevant signals, task-irrelevant VPL may occur if the feature is not weak.
Here, a dual plasticity model is proposed to comprehensively explain the apparently contradictory results and task-irrelevant VPL as well as task-relevant VPL. In this model, there are feature-based plasticity and task-based plasticity. Feature-based plasticity reflects a change in a representation of a feature presented during training, irrespective of whether the feature is task-relevant or task-irrelevant. Task-based plasticity reflects a change in processing of the task performed during training. Feature-based plasticity occurs in a lower stage than task-based plasticity and constrains VPL only to the trained location and feature. Task-based plasticity constrains VPL only to the trained task.
One possible reason for a variety of models of VPL is that different researchers tend to use different sets of parameters as their stimuli, tasks, etc in conducting their VPL studies. This practice has made it very difficult to make direct comparisons between the findings and to find, if any, general rules of VPL. Given a huge data set, timing is second to none to start efforts to build a comprehensive framework of VPL, rather than accepting only a portion of data in accord with a model.
Acknowledgments
We are grateful to Drs. Daniel Schacter, Cong Yu, and Lihan Chen for valuable comments on an early draft. This research has been supported by R01 AG031941, R01 MH091801, R01 EY015980 and R01 EY019466.
References
- Adab HZ, Vogels R. Practicing coarse orientation discrimination improves orientation signals in macaque cortical area v4. Current biology : CB. 2011;21:1661–6. doi: 10.1016/j.cub.2011.08.037. [DOI] [PubMed] [Google Scholar]
- Adini Y, Sagi D, Tsodyks M. Context-enabled learning in the human visual system. Nature. 2002;415:790–3. doi: 10.1038/415790a. [DOI] [PubMed] [Google Scholar]
- Ahissar M, Hochstein S. Attentional control of early perceptual learning. Proceedings of the National Academy of Sciences of the United States of America. 1993;90:5718–22. doi: 10.1073/pnas.90.12.5718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahissar M, Hochstein S. Task difficulty and the specificity of perceptual learning. Nature. 1997;387:401–6. doi: 10.1038/387401a0. [DOI] [PubMed] [Google Scholar]
- Andersen GJ. Aging and Vision: Changes in Function and Performance from Optics to Perception. Wiley Interdiscip Rev Cogn Sci. 2012;3:403–10. doi: 10.1002/wcs.1167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andersen GJ, Ni R, Bower JD, Watanabe T. Perceptual learning, aging, and improved visual performance in early stages of visual processing. Journal of vision. 2010;10:4–11. doi: 10.1167/10.13.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashby FG, Maddox WT. Human category learning 2.0. Annals of the New York Academy of Sciences. 2011;1224:147–61. doi: 10.1111/j.1749-6632.2010.05874.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baldassarre A, Lewis CM, Committeri G, Snyder AZ, Romani GL, Corbetta M. Individual variability in functional connectivity predicts performance of a perceptual task. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:3516–21. doi: 10.1073/pnas.1113148109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ball K, Sekuler R. Direction-specific improvement in motion discrimination. Vision research. 1987;27:953–65. doi: 10.1016/0042-6989(87)90011-3. [DOI] [PubMed] [Google Scholar]
- Barbot A, Landy MS, Carrasco M. Exogenous attention enhances 2nd-order contrast sensitivity. Vision research. 2011;51:1086–98. doi: 10.1016/j.visres.2011.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baumann O, Endestad T, Magnussen S, Greenlee MW. Delayed discrimination of spatial frequency for gratings of different orientation: behavioral and fMRI evidence for low-level perceptual memory stores in early visual cortex. Experimental brain research. 2008;188:363–9. doi: 10.1007/s00221-008-1366-0. [DOI] [PubMed] [Google Scholar]
- Bejjanki VR, Beck JM, Lu ZL, Pouget A. Perceptual learning as improved probabilistic inference in early sensory areas. Nat Neurosci. 2011;14:642–8. doi: 10.1038/nn.2796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett PJ, Sekuler AB, McIntosh AR, Della-Maggiore V. The effects of aging on visual memory: evidence for functional reorganization of cortical networks. Acta Psychol (Amst) 2001;107:249–73. doi: 10.1016/s0001-6918(01)00037-3. [DOI] [PubMed] [Google Scholar]
- Beste C, Dinse HR. Learning without training. Current biology : CB. 2013;23:R489–99. doi: 10.1016/j.cub.2013.04.044. [DOI] [PubMed] [Google Scholar]
- Beste C, Wascher E, Gunturkun O, Dinse HR. Improvement and impairment of visually guided behavior through LTP- and LTD-like exposure-based visual learning. Current biology : CB. 2011;21:876–82. doi: 10.1016/j.cub.2011.03.065. [DOI] [PubMed] [Google Scholar]
- Bi T, Chen J, Zhou T, He Y, Fang F. Function and structure of human left fusiform cortex are closely associated with perceptual learning of faces. Current biology : CB. 2014;24:222–7. doi: 10.1016/j.cub.2013.12.028. [DOI] [PubMed] [Google Scholar]
- Bouvier SE, Cardinal KS, Engel SA. Activity in visual area V4 correlates with surface perception. Journal of vision. 2008;8:28, 1–9. doi: 10.1167/8.7.28. [DOI] [PubMed] [Google Scholar]
- Bower JD, Watanabe T, Andersen GJ. Perceptual learning and aging: improved performance for low-contrast motion discrimination. Frontiers in psychology. 2013;4:66. doi: 10.3389/fpsyg.2013.00066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braddick OJ, O'Brien JM, Wattam-Bell J, Atkinson J, Hartley T, Turner R. Brain areas sensitive to coherent visual motion. Perception. 2001;30:61–72. doi: 10.1068/p3048. [DOI] [PubMed] [Google Scholar]
- Carrasco M, Rosenbaum A, Giordano A. Exogenous attention: Less effort, more learning! J of V. 2008;8:1095a. [Google Scholar]
- Choi H, Watanabe T. Is perceptual learning associated with changes in a sensory region? F1000 Biol Rep. 2012;4:B4–24. doi: 10.3410/B4-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chowdhury SA, DeAngelis GC. Fine discrimination training alters the causal contribution of macaque area MT to depth perception. Neuron. 2008;60:367–77. doi: 10.1016/j.neuron.2008.08.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crist RE, Kapadia MK, Westheimer G, Gilbert CD. Perceptual learning of spatial localization: specificity for orientation, position, and context. Journal of neurophysiology. 1997;78:2889–94. doi: 10.1152/jn.1997.78.6.2889. [DOI] [PubMed] [Google Scholar]
- Crist RE, Li W, Gilbert CD. Learning to see: experience and attention in primary visual cortex. Nat Neurosci. 2001;4:519–25. doi: 10.1038/87470. [DOI] [PubMed] [Google Scholar]
- de Villers-Sidani E, Merzenich MM. Lifelong plasticity in the rat auditory cortex: basic mechanisms and role of sensory experience. Progress in brain research. 2011;191:119–31. doi: 10.1016/B978-0-444-53752-2.00009-6. [DOI] [PubMed] [Google Scholar]
- Deloss DJ, Watanabe T, Andersen GJ. Optimization of perceptual learning: Effects of task difficulty and external noise in older adults. Vision research. 2013 doi: 10.1016/j.visres.2013.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dewald AD, Sinnett S, Doumas LA. Conditions of directed attention inhibit recognition performance for explicitly presented target-aligned irrelevant stimuli. Acta Psychol (Amst) 2011;138:60–7. doi: 10.1016/j.actpsy.2011.05.006. [DOI] [PubMed] [Google Scholar]
- Dewald AD, Sinnett S, Doumas LA. A window of perception when diverting attention? Enhancing recognition for explicitly presented, unattended, and irrelevant stimuli by target alignment. J Exp Psychol Hum Percept Perform. 2013;39:1304–12. doi: 10.1037/a0031210. [DOI] [PubMed] [Google Scholar]
- Dill M, Fahle M. The role of visual field position in pattern-discrimination learning. Proc R Soc Lond B Biol Sci. 1997;264:1031–6. doi: 10.1098/rspb.1997.0142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dosher BA, Jeter P, Liu J, Lu ZL. An integrated reweighting theory of perceptual learning. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:13678–83. doi: 10.1073/pnas.1312552110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dosher BA, Liu SH, Blair N, Lu ZL. The spatial window of the perceptual template and endogenous attention. Vision research. 2004;44:1257–71. doi: 10.1016/j.visres.2004.01.011. [DOI] [PubMed] [Google Scholar]
- Dosher BA, Lu ZL. Perceptual learning reflects external noise filtering and internal noise reduction through channel reweighting. Proceedings of the National Academy of Sciences of the United States of America. 1998;95:13988–93. doi: 10.1073/pnas.95.23.13988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dosher BA, Lu ZL. Mechanisms of perceptual learning. Vision research. 1999;39:3197–221. doi: 10.1016/s0042-6989(99)00059-0. [DOI] [PubMed] [Google Scholar]
- Eagleman D. Incognito: The Secret Lives of the Brain. Pantheon (US); Canongate (UK): 2011. [Google Scholar]
- Fahle M. Perceptual learning: gain without pain? Nat Neurosci. 2002;5:923–4. doi: 10.1038/nn1002-923. [DOI] [PubMed] [Google Scholar]
- Fahle M. Perceptual learning and sensomotor flexibility: cortical plasticity under attentional control? Philosophical transactions of the Royal Society of London Series B, Biological sciences. 2009;364:313–9. doi: 10.1098/rstb.2008.0267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fahle M, Edelman S. Long-term learning in vernier acuity: effects of stimulus orientation, range and of feedback. Vision research. 1993;33:397–412. doi: 10.1016/0042-6989(93)90094-d. [DOI] [PubMed] [Google Scholar]
- Fahle M, Edelman S, Poggio T. Fast perceptual learning in hyperacuity. Vision research. 1995;35:3003–13. doi: 10.1016/0042-6989(95)00044-z. [DOI] [PubMed] [Google Scholar]
- Fahle M, Poggio T. Perceptual learning. Cambridge, Mass: MIT Press; 2002. [Google Scholar]
- Felleman DJ, Van Essen DC. Distributed hierarchical processing in the primate cerebral cortex. Cereb Cortex. 1991;1:1–47. doi: 10.1093/cercor/1.1.1-a. [DOI] [PubMed] [Google Scholar]
- Fine I, Jacobs RA. Comparing perceptual learning tasks: a review. Journal of vision. 2002;2:190–203. doi: 10.1167/2.2.5. [DOI] [PubMed] [Google Scholar]
- Fiorentini A, Berardi N. Perceptual learning specific for orientation and spatial frequency. Nature. 1980;287:43–4. doi: 10.1038/287043a0. [DOI] [PubMed] [Google Scholar]
- Folstein JR, Gauthier I, Palmeri TJ. Mere exposure alters category learning of novel objects. Frontiers in psychology. 2010;1:40. doi: 10.3389/fpsyg.2010.00040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furmanski CS, Schluppeck D, Engel SA. Learning strengthens the response of primary visual cortex to simple patterns. Current biology : CB. 2004;14:573–8. doi: 10.1016/j.cub.2004.03.032. [DOI] [PubMed] [Google Scholar]
- Garrigan P, Kellman PJ. Perceptual learning depends on perceptual constancy. Proceedings of the National Academy of Sciences of the United States of America. 2008;105:2248–53. doi: 10.1073/pnas.0711878105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghose GM, Yang T, Maunsell JH. Physiological correlates of perceptual learning in monkey V1 and V2. Journal of neurophysiology. 2002;87:1867–88. doi: 10.1152/jn.00690.2001. [DOI] [PubMed] [Google Scholar]
- Gilbert CD. Early perceptual learning. Proceedings of the National Academy of Sciences of the United States of America. 1994;91:1195–7. doi: 10.1073/pnas.91.4.1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert CD, Li W. Adult visual cortical plasticity. Neuron. 2012;75:250–64. doi: 10.1016/j.neuron.2012.06.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gold JI, Law CT, Connolly P, Bennur S. The relative influences of priors and sensory evidence on an oculomotor decision variable during perceptual learning. Journal of neurophysiology. 2008;100:2653–68. doi: 10.1152/jn.90629.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gold JI, Law CT, Connolly P, Bennur S. Relationships between the threshold and slope of psychometric and neurometric functions during perceptual learning: implications for neuronal pooling. Journal of neurophysiology. 2009;103:140–54. doi: 10.1152/jn.00744.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gold JI, Watanabe T. Perceptual learning. Current biology : CB. 2010;20:R46–8. doi: 10.1016/j.cub.2009.10.066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstone RL. Perceptual learning. Annual Review of Psychology. 1998;49:585–612. doi: 10.1146/annurev.psych.49.1.585. [DOI] [PubMed] [Google Scholar]
- Gu Y, Liu S, Fetsch CR, Yang Y, Fok S, et al. Perceptual learning reduces interneuronal correlations in macaque visual cortex. Neuron. 2012;71:750–61. doi: 10.1016/j.neuron.2011.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutnisky DA, Hansen BJ, Iliescu BF, Dragoi V. Attention Alters Visual Plasticity during Exposure-Based Learning. Current biology : CB. 2009 doi: 10.1016/j.cub.2009.01.063. [DOI] [PubMed] [Google Scholar]
- Hammer R, Sloutsky V, Grill-Spector K. CogSci2012. Sapporo, Japan: Curran Associates, Inc.; 2012. paper 0084. [Google Scholar]
- Harris H, Gliksberg M, Sagi D. Generalized Perceptual Learning in the Absence of Sensory Adaptation. Current biology : CB. 2012 doi: 10.1016/j.cub.2012.07.059. [DOI] [PubMed] [Google Scholar]
- Harris JP, Fahle M. The detection and discrimination of spatial offsets. Vision research. 1995;35:51–8. doi: 10.1016/0042-6989(94)e0082-v. [DOI] [PubMed] [Google Scholar]
- Heekeren HR, Marrett S, Bandettini PA, Ungerleider LG. A general mechanism for perceptual decision-making in the human brain. Nature. 2004;431:859–62. doi: 10.1038/nature02966. [DOI] [PubMed] [Google Scholar]
- Hensch TK. Critical period mechanisms in developing visual cortex. Curr Top Dev Biol. 2005a;69:215–37. doi: 10.1016/S0070-2153(05)69008-4. [DOI] [PubMed] [Google Scholar]
- Hensch TK. Critical period plasticity in local cortical circuits. Nat Rev Neurosci. 2005b;6:877–88. doi: 10.1038/nrn1787. [DOI] [PubMed] [Google Scholar]
- Hochstein S, Ahissar M. View from the top: hierarchies and reverse hierarchies in the visual system. Neuron. 2002;36:791–804. doi: 10.1016/s0896-6273(02)01091-7. [DOI] [PubMed] [Google Scholar]
- Hua T, Bao P, Huang CB, Wang Z, Xu J, et al. Perceptual learning improves contrast sensitivity of V1 neurons in cats. Current biology : CB. 2010;20:887–94. doi: 10.1016/j.cub.2010.03.066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang X, Lu H, Tjan BS, Zhou Y, Liu Z. Motion perceptual learning: when only task-relevant information is learned. Journal of vision. 2007;7:14, 1–10. doi: 10.1167/7.10.14. [DOI] [PubMed] [Google Scholar]
- Hubel DH, Wiesel TN. Effects of Monocular Deprivation in Kittens. Naunyn Schmiedebergs Arch Pharmacol. 1964;248:492–7. doi: 10.1007/BF00348878. [DOI] [PubMed] [Google Scholar]
- Hubel DH, Wiesel TN. Receptive fields and functional architecture of monkey striate cortex. J Physiol. 1968;195:215–43. doi: 10.1113/jphysiol.1968.sp008455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kahnt T, Grueschow M, Speck O, Haynes JD. Perceptual learning and decision-making in human medial frontal cortex. Neuron. 2011;70:549–59. doi: 10.1016/j.neuron.2011.02.054. [DOI] [PubMed] [Google Scholar]
- Kamitani Y, Tong F. Decoding the visual and subjective contents of the human brain. Nat Neurosci. 2005;8:679–85. doi: 10.1038/nn1444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karmarkar UR, Dan Y. Experience-dependent plasticity in adult visual cortex. Neuron. 2006;52:577–85. doi: 10.1016/j.neuron.2006.11.001. [DOI] [PubMed] [Google Scholar]
- Karni A, Sagi D. Where practice makes perfect in texture discrimination: evidence for primary visual cortex plasticity. Proceedings of the National Academy of Sciences of the United States of America. 1991;88:4966–70. doi: 10.1073/pnas.88.11.4966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karni A, Sagi D. The time course of learning a visual skill. Nature. 1993;365:250–2. doi: 10.1038/365250a0. [DOI] [PubMed] [Google Scholar]
- Klein SA. 2007 [Google Scholar]
- Koyama S, Harner A, Watanabe T. Task-dependent changes of the psychophysical motion-tuning functions in the course of perceptual learning. Perception. 2004;33:1139–47. doi: 10.1068/p5195. [DOI] [PubMed] [Google Scholar]
- Koyama S, Sasaki Y, Andersen GJ, Tootell RB, Matsuura M, Watanabe T. Separate processing of different global-motion structures in visual cortex is revealed by FMRI. Current biology : CB. 2005;15:2027–32. doi: 10.1016/j.cub.2005.10.069. [DOI] [PubMed] [Google Scholar]
- Law CT, Gold JI. Neural correlates of perceptual learning in a sensory-motor, but not a sensory, cortical area. Nat Neurosci. 2008;11:505–13. doi: 10.1038/nn2070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leclercq V, Le Dantec CC, Seitz AR. Encoding of episodic information through fast task-irrelevant perceptual learning. Vision research. 2013 doi: 10.1016/j.visres.2013.09.006. [DOI] [PubMed] [Google Scholar]
- Leclercq V, Seitz AR. The impact of orienting attention in fast task-irrelevant perceptual learning. Attention, perception & psychophysics. 2012;74:648–60. doi: 10.3758/s13414-012-0270-7. [DOI] [PubMed] [Google Scholar]
- Lee SH, Kwan AC, Zhang S, Phoumthipphavong V, Flannery JG, et al. Activation of specific interneurons improves V1 feature selectivity and visual perception. Nature. 2012;488:379–83. doi: 10.1038/nature11312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levi DM. Prentice award lecture 2011: removing the brakes on plasticity in the amblyopic brain. Optometry and vision science : official publication of the American Academy of Optometry. 2012;89:827–38. doi: 10.1097/OPX.0b013e318257a187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levi DM, Li RW. Improving the performance of the amblyopic visual system. Philosophical transactions of the Royal Society of London Series B, Biological sciences. 2009a;364:399–407. doi: 10.1098/rstb.2008.0203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levi DM, Li RW. Perceptual learning as a potential treatment for amblyopia: a mini-review. Vision research. 2009b;49:2535–49. doi: 10.1016/j.visres.2009.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis CM, Baldassarre A, Committeri G, Romani GL, Corbetta M. Learning sculpts the spontaneous activity of the resting human brain. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:17558–63. doi: 10.1073/pnas.0902455106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J, Thompson B, Deng D, Chan LY, Yu M, Hess RF. Dichoptic training enables the adult amblyopic brain to learn. Current biology : CB. 2013;23:R308–9. doi: 10.1016/j.cub.2013.01.059. [DOI] [PubMed] [Google Scholar]
- Li N, DiCarlo JJ. Unsupervised natural experience rapidly alters invariant object representation in visual cortex. Science. 2008;321:1502–7. doi: 10.1126/science.1160028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li N, Dicarlo JJ. Neuronal learning of invariant object representation in the ventral visual stream is not dependent on reward. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2012;32:6611–20. doi: 10.1523/JNEUROSCI.3786-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu J, Lu ZL, Dosher BA. Augmented Hebbian reweighting: interactions between feedback and training accuracy in perceptual learning. Journal of vision. 2010;10:29. doi: 10.1167/10.10.29. [DOI] [PubMed] [Google Scholar]
- Liu Z. Perceptual learning in motion discrimination that generalizes across motion directions. Proceedings of the National Academy of Sciences of the United States of America. 1999;96:14085–7. doi: 10.1073/pnas.96.24.14085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Logothetis NK, Pauls J, Augath M, Trinath T, Oeltermann A. Neurophysiological investigation of the basis of the fMRI signal. Nature. 2001;412:150–7. doi: 10.1038/35084005. [DOI] [PubMed] [Google Scholar]
- Lu ZL, Chu W, Dosher BA, Lee S. Independent perceptual learning in monocular and binocular motion systems. Proceedings of the National Academy of Sciences of the United States of America. 2005;102:5624–9. doi: 10.1073/pnas.0501387102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu ZL, Hua T, Huang CB, Zhou Y, Dosher BA. Visual perceptual learning. Neurobiol Learn Mem. 2011;95:145–51. doi: 10.1016/j.nlm.2010.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKee SP, Westheimer G. Improvement in vernier acuity with practice. Percept Psychophys. 1978;24:258–62. doi: 10.3758/bf03206097. [DOI] [PubMed] [Google Scholar]
- Morishita H, Hensch TK. Critical period revisited: impact on vision. Current opinion in neurobiology. 2008;18:101–7. doi: 10.1016/j.conb.2008.05.009. [DOI] [PubMed] [Google Scholar]
- Movshon J, Adelson E, Gizzi M, Newsome W. The analysis of moving visual patterns. Pattern recognition mechanisms. 1985;54:117–51. [Google Scholar]
- Newsome WT, Pare EB. A selective impairment of motion perception following lesions of the middle temporal visual area (MT) The Journal of neuroscience : the official journal of the Society for Neuroscience. 1988;8:2201–11. doi: 10.1523/JNEUROSCI.08-06-02201.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ooi TL, Su YR, Natale DM, He ZJ. A push-pull treatment for strengthening the 'lazy eye' in amblyopia. Current biology : CB. 2013;23:R309–10. doi: 10.1016/j.cub.2013.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Op de Beeck HP, Baker CI. The neural basis of visual object learning. Trends in cognitive sciences. 2010;14:22–30. doi: 10.1016/j.tics.2009.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrov AA, Dosher BA, Lu ZL. The dynamics of perceptual learning: an incremental reweighting model. Psychol Rev. 2005;112:715–43. doi: 10.1037/0033-295X.112.4.715. [DOI] [PubMed] [Google Scholar]
- Petrov AA, Dosher BA, Lu ZL. Perceptual learning without feedback in non-stationary contexts: data and model. Vision research. 2006;46:3177–97. doi: 10.1016/j.visres.2006.03.022. [DOI] [PubMed] [Google Scholar]
- Piech V, Li W, Reeke GN, Gilbert CD. Network model of top-down influences on local gain and contextual interactions in visual cortex. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:E4108–17. doi: 10.1073/pnas.1317019110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poggio T, Fahle M, Edelman S. Fast perceptual learning in visual hyperacuity. Science. 1992;256:1018–21. doi: 10.1126/science.1589770. [DOI] [PubMed] [Google Scholar]
- Polat U, Ma-Naim T, Belkin M, Sagi D. Improving vision in adult amblyopia by perceptual learning. Proceedings of the National Academy of Sciences of the United States of America. 2004;101:6692–7. doi: 10.1073/pnas.0401200101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reed A, Riley J, Carraway R, Carrasco A, Perez C, et al. Cortical map plasticity improves learning but is not necessary for improved performance. Neuron. 2011;70:121–31. doi: 10.1016/j.neuron.2011.02.038. [DOI] [PubMed] [Google Scholar]
- Rees G, Friston K, Koch C. A direct quantitative relationship between the functional properties of human and macaque V5. Nat Neurosci. 2000;3:716–23. doi: 10.1038/76673. [DOI] [PubMed] [Google Scholar]
- Roe AW, Ts'o DY. Visual topography in primate V2: multiple representation across functional stripes. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1995;15:3689–715. doi: 10.1523/JNEUROSCI.15-05-03689.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenthal O, Humphreys GW. Perceptual organization without perception. The subliminal learning of global contour. Psychol Sci. 2010;21:1751–8. doi: 10.1177/0956797610389188. [DOI] [PubMed] [Google Scholar]
- Saarinen J, Levi DM. Perceptual learning in vernier acuity: what is learned? Vision research. 1995;35:519–27. doi: 10.1016/0042-6989(94)00141-8. [DOI] [PubMed] [Google Scholar]
- Sagi D. Perceptual learning in Vision Research. Vision research. 2011;51:1552–66. doi: 10.1016/j.visres.2010.10.019. [DOI] [PubMed] [Google Scholar]
- Sagi D, Tanne D. Perceptual learning: learning to see. Current opinion in neurobiology. 1994;4:195–9. doi: 10.1016/0959-4388(94)90072-8. [DOI] [PubMed] [Google Scholar]
- Sasaki S, Watanabe T. Perceptual Learning. In: Werner J, Chalupa L, editors. VIsual Neurosceinces. Cambridge: MIT Press; 2012. pp. 991–1000. [Google Scholar]
- Sasaki Y, Nanez JE, Watanabe T. Advances in visual perceptual learning and plasticity. Nat Rev Neurosci. 2010;11:53–60. doi: 10.1038/nrn2737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoups A, Vogels R, Qian N, Orban G. Practising orientation identification improves orientation coding in V1 neurons. Nature. 2001;412:549–53. doi: 10.1038/35087601. [DOI] [PubMed] [Google Scholar]
- Schoups AA, Orban GA. Interocular transfer in perceptual learning of a pop-out discrimination task. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:7358–62. doi: 10.1073/pnas.93.14.7358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoups AA, Vogels R, Orban GA. Human perceptual learning in identifying the oblique orientation: retinotopy, orientation specificity and monocularity. J Physiol. 1995;483(Pt 3):797–810. doi: 10.1113/jphysiol.1995.sp020623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz S, Maquet P, Frith C. Neural correlates of perceptual learning: a functional MRI study of visual texture discrimination. Proceedings of the National Academy of Sciences of the United States of America. 2002;99:17137–42. doi: 10.1073/pnas.242414599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seitz A, Kim D, Watanabe T. Rewards evoke learning of unconsciously processed visual stimuli in adult humans. Neuron. 2009;61:700–07. doi: 10.1016/j.neuron.2009.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seitz A, Watanabe T. A unified model for perceptual learning. Trends in cognitive sciences. 2005;9:329–34. doi: 10.1016/j.tics.2005.05.010. [DOI] [PubMed] [Google Scholar]
- Seitz AR, Dinse HR. A common framework for perceptual learning. Current opinion in neurobiology. 2007;17:148–53. doi: 10.1016/j.conb.2007.02.004. [DOI] [PubMed] [Google Scholar]
- Seitz AR, Lefebvre C, Watanabe T, Jolicoeur P. Requirement for high-level processing in subliminal learning. Current biology : CB. 2005a;15:R753–5. doi: 10.1016/j.cub.2005.09.009. [DOI] [PubMed] [Google Scholar]
- Seitz AR, Watanabe T. Psychophysics: Is subliminal learning really passive? Nature. 2003;422:36. doi: 10.1038/422036a. [DOI] [PubMed] [Google Scholar]
- Seitz AR, Yamagishi N, Werner B, Goda N, Kawato M, Watanabe T. Task-specific disruption of perceptual learning. Proceedings of the National Academy of Sciences of the United States of America. 2005b;102:14895–900. doi: 10.1073/pnas.0505765102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shadlen MN. 2014 [Google Scholar]
- Shadlen MN, Newsome WT. Motion perception: seeing and deciding. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:628–33. doi: 10.1073/pnas.93.2.628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shadlen MN, Newsome WT. Neural basis of a perceptual decision in the parietal cortex (area LIP) of the rhesus monkey. Journal of neurophysiology. 2001;86:1916–36. doi: 10.1152/jn.2001.86.4.1916. [DOI] [PubMed] [Google Scholar]
- Shibata K, Chang LH, Kim D, Nanez JE, Sr, Kamitani Y, et al. Decoding Reveals Plasticity in V3A as a Result of Motion Perceptual Learning. PLoS One. 2012a;7:e44003. doi: 10.1371/journal.pone.0044003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shibata K, Kawato M, Sasaki S, Watanabe T. Monocular deprivation boosts long-term visual plasticity. Current biology : CB. 2012b;22:R291–2. doi: 10.1016/j.cub.2012.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shibata K, Sasaki Y, Kawato M, Watanabe T. Perceptual learning is associated with different types of plasticity. Journal of vision. 2013;13:604. [Google Scholar]
- Shibata K, Watanabe T. Preference suppression caused by misattribution of task-irrelevant subliminal motion. Royal Society of London (B) 2012;279:3443–8. doi: 10.1098/rspb.2012.0797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shibata K, Watanabe T, Sasaki Y, Kawato M. Perceptual learning incepted by decoded fMRI neurofeedback without stimulus presentation. Science. 2011;334:1413–5. doi: 10.1126/science.1212003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiu LP, Pashler H. Improvement in line orientation discrimination is retinally local but dependent on cognitive set. Percept Psychophys. 1992;52:582–8. doi: 10.3758/bf03206720. [DOI] [PubMed] [Google Scholar]
- Tootell RB, Mendola JD, Hadjikhani NK, Ledden PJ, Liu AK, et al. Functional analysis of V3A and related areas in human visual cortex. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1997;17:7060–78. doi: 10.1523/JNEUROSCI.17-18-07060.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ts'o DY, Gilbert CD. The organization of chromatic and spatial interactions in the primate striate cortex. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1988;8:1712–27. doi: 10.1523/JNEUROSCI.08-05-01712.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsushima Y, Sasaki Y, Watanabe T. Greater disruption due to failure of inhibitory control on an ambiguous distractor. Science. 2006;314:1786–8. doi: 10.1126/science.1133197. [DOI] [PubMed] [Google Scholar]
- Tsushima Y, Seitz A, Watanabe T. Task-irrelevant learning occurs only when the irrelevant feature is weak. Current Biol. 2008;18:R516–7. doi: 10.1016/j.cub.2008.04.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsushima Y, Watanabe T. Roles of attention in perceptual learning from perspectives of psychophysics and animal learning. Learn Behav. 2009;37:126–32. doi: 10.3758/LB.37.2.126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaina LM, Belliveau JW, des Roziers EB, Zeffiro TA. Neural systems underlying learning and representation of global motion. Proceedings of the National Academy of Sciences of the United States of America. 1998;95:12657–62. doi: 10.1073/pnas.95.21.12657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vlahou EL, Protopapas A, Seitz AR. Implicit training of nonnative speech stimuli. J Exp Psychol Gen. 2012;141:363–81. doi: 10.1037/a0025014. [DOI] [PubMed] [Google Scholar]
- Walker MP, Stickgold R, Alsop D, Gaab N, Schlaug G. Sleep-dependent motor memory plasticity in the human brain. Neuroscience. 2005;133:911–7. doi: 10.1016/j.neuroscience.2005.04.007. [DOI] [PubMed] [Google Scholar]
- Wang R, Zhang JY, Klein SA, Levi DM, Yu C. Task relevancy and demand modulate double-training enabled transfer of perceptual learning. Vision research. 2012;61:33–8. doi: 10.1016/j.visres.2011.07.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watamaniuk SN, Sekuler R, Williams DW. Direction perception in complex dynamic displays: the integration of direction information. Vision research. 1989;29:47–59. doi: 10.1016/0042-6989(89)90173-9. [DOI] [PubMed] [Google Scholar]
- Watanabe T, Nanez JE, Sasaki Y. Perceptual learning without perception. Nature. 2001;413:844–8. doi: 10.1038/35101601. [DOI] [PubMed] [Google Scholar]
- Watanabe T, Nanez JE, Sr, Koyama S, Mukai I, Liederman J, Sasaki Y. Greater plasticity in lower-level than higher-level visual motion processing in a passive perceptual learning task. Nat Neurosci. 2002;5:1003–9. doi: 10.1038/nn915. [DOI] [PubMed] [Google Scholar]
- Westheimer G, Crist RE, Gorski L, Gilbert CD. Configuration specificity in bisection acuity. Vision research. 2001;41:1133–8. doi: 10.1016/s0042-6989(00)00320-5. [DOI] [PubMed] [Google Scholar]
- Witthoft N, Davidenko N, Grill-Spector K. CogSci2009. Austin, TX: Curran Associates, Inc.; 2009. pp. 3058–63. [Google Scholar]
- Xiao LQ, Zhang JY, Wang R, Klein SA, Levi DM, Yu C. Complete transfer of perceptual learning across retinal locations enabled by double training. Current biology : CB. 2008;18:1922–6. doi: 10.1016/j.cub.2008.10.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu JP, He ZJ, Ooi TL. Effectively reducing sensory eye dominance with a push-pull perceptual learning protocol. Current biology : CB. 2010;20:1864–8. doi: 10.1016/j.cub.2010.09.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu JP, He ZJ, Ooi TL. Further support for the importance of the suppressive signal (pull) during the push-pull perceptual training. Vision research. 2012a;61:60–9. doi: 10.1016/j.visres.2012.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu JP, He ZJ, Ooi TL. Perceptual learning to reduce sensory eye dominance beyond the focus of top-down visual attention. Vision research. 2012b;61:39–47. doi: 10.1016/j.visres.2011.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu JP, He ZJ, Ooi TL. Push-pull training reduces foveal sensory eye dominance within the early visual channels. Vision research. 2012c;61:48–59. doi: 10.1016/j.visres.2011.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang T, Maunsell JH. The effect of perceptual learning on neuronal responses in monkey visual area V4. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2004;24:1617–26. doi: 10.1523/JNEUROSCI.4442-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yotsumoto Y, Sasaki Y, Chan P, Vasios C, Bonmassar G, et al. Location-specific cortical activation changes during sleep after training for perceptual learning. Current biology : CB. 2009;19:1278–82. doi: 10.1016/j.cub.2009.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yotsumoto Y, Watanabe T, Sasaki Y. Different dynamics of performance and brain activation in the time course of perceptual learning. Neuron. 2008;57:827–33. doi: 10.1016/j.neuron.2008.02.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young KG, Li RW, Levi DM, Klein SA, Huang EY. Invest Ophthalmol Vis Sci. 2004;4363 [Google Scholar]
- Zeki S. A vision of the brain. Oxford: Blackwell Scientific Publications; 1993. [Google Scholar]
- Zhang J, Kourtzi Z. Learning-dependent plasticity with and without training in the human brain. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:13503–8. doi: 10.1073/pnas.1002506107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang JY, Kuai SG, Xiao LQ, Klein SA, Levi DM, Yu C. Stimulus coding rules for perceptual learning. PLoS Biol. 2008;6:e197. doi: 10.1371/journal.pbio.0060197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang JY, Zhang GL, Xiao LQ, Klein SA, Levi DM, Yu C. Rule-based learning explains visual perceptual learning and its specificity and transfer. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2010a;30:12323–8. doi: 10.1523/JNEUROSCI.0704-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang T, Xiao LQ, Klein SA, Levi DM, Yu C. Decoupling location specificity from perceptual learning of orientation discrimination. Vision research. 2010b;50:368–74. doi: 10.1016/j.visres.2009.08.024. [DOI] [PubMed] [Google Scholar]